I have just come across a very interesting paper by Akshay Nag and colleagues at the Institute of Himalayan Bioresource Technology, about Podophyllum hexandrum (Himalayan mayapple).

Himalayan mayapple, Podophyllum hexandrum. Image: Potterton’s nursery
The mayapple, an unlikely-looking relative of Berberis and Mahonia, is an important medicinal plant, traditionally used by local healers for ulcers, cuts and skin diseases. Found at altitudes of 3000 – 4200 m throughout the Himalayas, it is now endangered precisely because of its medicinal value. The plant’s roots contain chemicals, including podophyllotoxin, used in the manufacture of anticancer drugs; podophyllotoxin inhibits the assembly of microtubules, which both give cells their internal structure and are crucial for cell division. The Himalayan mayapple contains much larger quantities of podophyllotoxin than other sources, such as its North American sister species, Podophyllum peltatum, though absolute concentrations vary with location, season and with the age of the plant. Over-exploitation, together with habitat fragmentation, overgrazing and slow natural regeneration mean P. podophyllum is now an endangered species. To add to the problem, Himalayan mayapple has a large genome (16 Gb) but a small number of chromosomes (only six pairs), which is likely to reduce its ability to adapt to a changing climate (Knight et al., 2005).
Most plants of the Berberidaceae or barberry family, including P. peltatum are self-incompatible (SI). This means that pollen from a given plant cannot fertilise the stigma of the same plant – pollen from another (genetically different) individual is required. Such cross pollination is the default option for many species because it is the easiest way of maintaining all-important genetic diversity in the population. However this relies on there being sufficient pollinators available and usually means the plant has to invest in producing nectar, as a reward to attract said pollinators. Think of the Mahonia in your garden – an important early nectar source for the bees, wasps and hoverflies which pollinate it.

Wasp in search of an open Mahonia flower for its nectar reward. Image: urbanpollinators.blogspot.co.uk
P. hexandrum, however, depends largely on self-pollination – it is a self-compatible (SC) plant. The interesting thing to someone interested in plant evolution is that P. hexandrum is known to have diverged from its sister species, P. peltanum, around 6.5 million years ago, in the late Miocene, when the Himalayas were being lifted up and its current, alpine, habitat was first created. The theory goes that a shortage of pollinating insects in this new, harsher habitat, forced the early-flowering plant to adapt and evolve to be able to self-pollinate efficiently when necessary, in order to survive.
Self-compatible plants often have a higher level of biological fitness (i.e. are more likely to produce successful offspring) than self-incompatible ones when pollinators are in short supply, despite the potential risks of a reduced genetic pool (so called inbreeding depression). In American mayapple, seed set is very low when it is manually self-pollinated; inbreeding depression causes many of the fertilised ovules to abort. However Xiong et al. (2013) showed that Himalayan mayapple yielded a similarly high level of seed set (nearly 90 %) whether it underwent self or cross pollination. It seems likely that forms of the original mayapple which were able to successfully self-pollinate and produce offspring would have spread in the areas of the Himalayas which were rising, causing a transition from SI to SC as the norm. Eventually, a new species was born. This pattern, where the loss of pollinators drives the shift of breeding systems from SI to SC fits with Baker’s law, which proposes that species capable of ‘uniparental reproduction’ are more likely to establish themselves in a new habitat after long-distance dispersal than species which rely on finding a suitable pollinator (Baker, 1955). If the new colony is well adapted to its new environment, it will be able to spread throughout this area despite its limited capacity for genetic variation, with some traits becoming fixed because of genetic drift.
Whilst the seeds of a fertilised mayapple flower develop, the immature fruit is hidden by the large, drooping leaves, perhaps to protect it from unwanted attention by herbivores. However once the seeds are mature, the red, fleshy fruits are visible and attractive to Himalayan grazing animals and bords. These travel over great distances and so can disperse the seeds, safe in their hard seed coat, widely. One consequence of this, and of self-pollination, is that there are now many geographically-distinct populations of P. hexandrum within the Himalayan region, but just how genetically diverse are these?
Nag and his colleagues sampled 209 individual plants from 24 geographically-isolated populations in India alone. They assessed the genetic variability of P. hexandrum within these populations using amplified fragment length polymorphism (AFLP) markers, with a view to using this information to both inform conservation strategies and predict how the plant will respond to changing climatic conditions in the region. What they found was that all the individuals fitted into two major groups or genetic populations but that these two genetic populations were intermixed, in each location, with significant gene flow between them – the result of widespread seed dispersal by herbivores and birds. The red and green proportions of the pie charts in Fig.1 (below) indicate the proportion of each genetic population in the areas sampled. In some areas, largely Zanskar, Sikkim and the Pir Panjal range, the ‘red’ population predominates, whereas in other areas the ‘green’ one does.
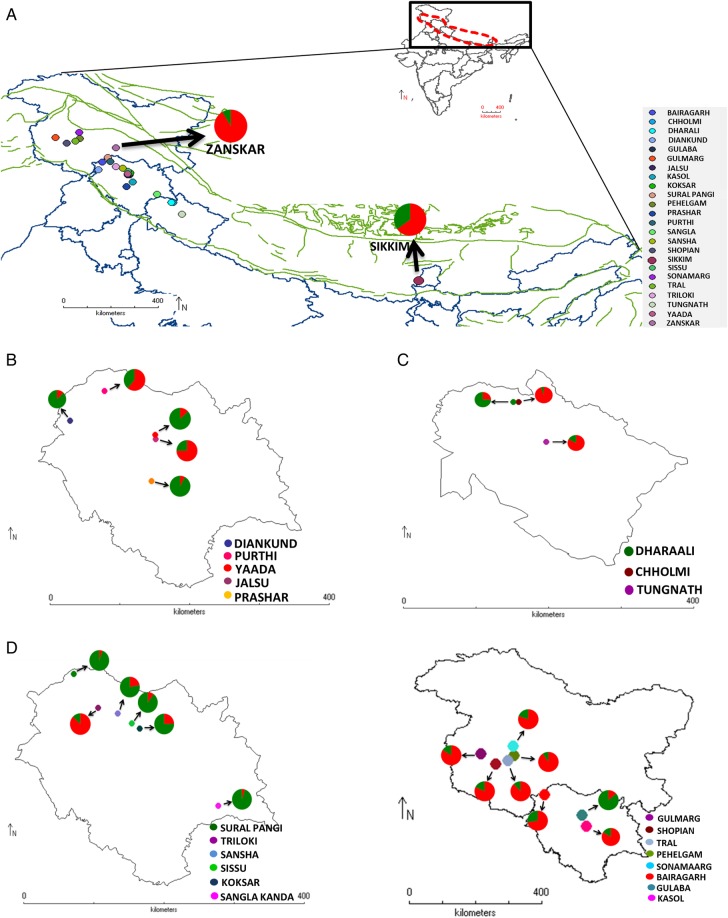
Fig. 1: Geographic distribution of sampled populations of P. hexandrum from the Indian Himalayas with pie charts representing the percentage of the two genetic pools from each of the populations. (A) Map representing all the sampled locations, (B) Dhauladhar range, (C) Shivalik/Garhwal Himalayas, (D) Greater Himalayas, (E) Pir Panjal range. (Nag et al., 2014)
The suggestion is that either all the populations in the Indian Himalayas are remnants of a single ancient population, once more widespread, or that they originated from two distinct genetic populations which once co-existed but then separated as a result of long distance dispersal and natural selection in response to their specific environments.
——–
Baker H.G. (1955). Self-compatibility and establishment after ‘longdistance’ dispersal. Evolution, 9,347–349.
Knight C.A., Molinari N.A. & Petrov D.A. (2005). The large genome constraint hypothesis: evolution, ecology and phenotype. Annals of Botany, 95, 177-190.
Nag A., Ahuja P.S. & Sharma R.K. (2014). Genetic diversity of high-elevation populations of an endangered medicinal plant. AoB Plants, 7, plu076.
Xiong Y-Z., Fang Q. & Huang S-Q. (2013). Pollinator scarcity drives the shift to delayed selfing in Himalayan mayapple, Podophyllum hexandrum (Berberidaceae). AoB Plants, 5, plt037.

really interesting Heather
I thought so, Sue.
[…] and Ladakh this summer is how plants are pollinated at different altitudes. I mentioned in ‘A case study in evolution – Podophyllum hexandrum’ that pollinating insects are in short supply at higher altitudes, forcing some plants to rely on […]
[…] of a sweet reward in the form of nectar. The bee and wasp-pollinated Mahonia described in ‘A case study in evolution – Podophyllum hexandrum’ is a case in point. A large range of different types of flowers are pollinated by bees and […]
[…] of centimeters tall studded the ground and I was thrilled to find Podophyllum hexandrum (see ‘A case study in evolution – Podophyllum hexandrum‘). Although much of the literature states that the main pressure on this endangered species […]
[…] With less than a week to go, it really feels like time to get back to thinking about the plants we’ll see in India. One thing we’d hoped to do last year, but didn’t really have time for, was to look at how the way in which plants are pollinated varies with factors such as altitude (see A case study in plant evolution). […]
Great reading article!
Glad you found it interesting 🙂
[…] tip of each branch and shrubs such as Viburnum, Daphne and Mahonia are sending out their scents to attract early insect pollinators. All are important sources of pollen and nectar for any early flying Honeybees and Bumblebees, […]
[…] My last blog post seems to have struck a chord with a number of people so I thought maybe I should post some weekly highlights of my daily attempts to notice nature more. Some days this week were easy, like the amble round the Botanic gardens on Thursday with my like-minded friend Heather, when the sun suddenly appeared after we’d had lunch. That was a multisensory feast, with sunshine pouring through fresh green bamboo leaves, swathes of snowdrops on sunny banks, birds singing their hearts out and the scent of early flowering shrubs in the winter garden. These early flowering shrubs are not just a delight for us but an important source of nectar and pollen for the earliest emerging insects. […]
[…] them or hate them (I love them) they are a very welcome source of food for early insects, including the bees we worry so much about. On my walk up through Cardiac Hill to the department I spotted the first […]